classification des êtres vivants
Classer les êtres vivants c’est rechercher leurs
points communs pour effectuer des regroupements, c’est rechercher une
certaine unité dans la diversité. Des résultats de cette activité
naît la notion de parenté entre les êtres vivants : plus des
êtres vivants présentent des caractères (appelés également
" attributs ") en commun, plus ils sont apparentés.
On devine derrière ces " parentés " une idée d’évolution.
 Classer selon certains critères : homologie et analogie
Classer selon certains critères : homologie et analogie
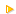 Il existe un obstacle majeur aux parentés ainsi trouvées. Les liens de
parenté ainsi établis reposent intégralement sur le choix des
critères de comparaison. Sont-ils tous fiables ?
Il existe un obstacle majeur aux parentés ainsi trouvées. Les liens de
parenté ainsi établis reposent intégralement sur le choix des
critères de comparaison. Sont-ils tous fiables ?
Si le critère principal choisi est par exemple le
milieu de vie : requin et dauphin sont les plus apparentés, ils
partagent ce milieu de vie en commun.
Si " l’allure " général des
animaux est comparé : requin et dauphin sont les plus apparentés,
ils ont une morphologie semblable, des nageoires…
Si le critère retenu est, soit l’appareil
respiratoire, soit le mode de reproduction (et les organes
associés) : alors le dauphin et le chien sont les plus
apparentés.
Il faut bien catégoriser les critères de
comparaison.
Tout d’abord, on ne peut comparer que ce qui est
réellement observé : les liens de parenté sont établis sur la
base des ressemblances et non des différences. Dire que deux êtres
vivants présentant de nombreuses ressemblances sont très apparentés
est probablement juste, par contre dire que deux êtres vivants
présentant peu de ressemblances (ou beaucoup de différences) sont peu
apparentés est faux. Les parentés ne peuvent s’établir qu’à
partir de trois êtres vivants dont deux seront plus apparentés
entre eux qu’avec le troisième, car ils présentent davantage de
caractères en commun. Dans le même esprit, c’est bien le partage de
caractères existant en commun qui est pris en compte et non pas l’absence
de caractère. Ainsi on ne décrit plus un groupe d’êtres vivants par
les caractères qu’il n’ont pas mais par ceux qu’ils ont et
partagent avec d’autres (on peut dire qu’il existe des Vertébrés,
ce sont les animaux qui possèdent un squelette axial interne formé de
vertèbres, mais on ne peut plus parler " d’Invertébrés "
dans les classifications actuelles).
Ensuite le milieu de vie ne permet pas d’effectuer
des liens de parenté : le monde vivant regorge d’exemples
montrant des êtres vivants fortement apparentés vivant dans des
environnements très différents (même chose concernant des individus d’une
même espèce) et inversement, des êtres vivants très différents
partageant les mêmes environnements.
Des morphologies particulières observées résultent
d’un " tri sélectif ", conséquence de l’action
de l’environnement sur les êtres vivants (voir la partie sur la
sélection naturelle) : la convergence de morphologie entre le
dauphin et le requin est le résultat de deux histoires évolutives
différentes mais concernant deux animaux dont la morphologie est
adaptée à la nage. Par contre, si on regarde de plus près leur mode
de locomotion, on remarque que le requin a effectivement des nageoires
constituées de rayons cartilagineux alors que le dauphin a des pattes
constituées de doigts osseux (comme nous) mais rapprochés les uns des
autres et recouverts par un épiderme qui n’individualise pas ces
doigts. Ces caractères morphologiques qui se ressemblent du fait de l’environnement
sont nommés caractères analogues. Les caractères analogues sont
associés à des fonctions et ne reflètent pas toujours des liens de
parenté. Ces caractères ne peuvent donc pas être pris en compte pour
effectuer des regroupements et donc des classements ou classifications
évolutives.
Nous utiliserons donc ce que l’on appelle des caractères homologues
pour effectuer des comparaisons. Ces caractères homologues ont tout d’abord
été des caractères provenant de comparaisons anatomiques (et non
morphologiques) ; on effectue actuellement des comparaison de
molécules homologues.
|
Si on compare la nageoire du requin et les pattes du
dauphin ainsi que du chien :
D’un point de vue morphologie (forme générale),
ce sont des caractères analogues (les formes sont en relation avec l’adaptation
au milieu de vie)
D’un point de vue anatomique (organisation
interne), ce sont de caractères homologues (les formes semblables sont
en relation avec les parentés)
Ci-contre : squelettes de membres
antérieur de chien et de dauphin. Le requin ne possède pas d'os dans
sa nageoire.
|
Comment savoir si les critères choisis pour classer des êtres vivants
correspondent à des caractères homologues et non analogues ?
Activité : effectuer des regroupements
Pour effectuer des regroupements d'êtres
vivants selon leurs parentés on réalise des groupes emboîtés
construits à partir des attributs (ce qui veut bien dire "ce que
les êtres vivants ont") qu'ils possèdent en communs. Le terme
"attribut" désigne aussi bien un caractère (qui sera
présent ou absent : donc deux possibilités) qu'un état de caractère
(qui lui peut présenter plusieurs variantes).
|
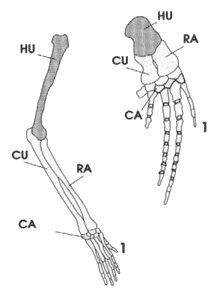
Par exemple, les six animaux du tableau ci contre
vont être décrits selon 4 attributs. L'observation permet d'établir
un tableau comparatif dans lequel "1" indique la
présence du caractère considéré et "0" indique
l'absence de ce caractère.
Remarque : les 4 attributs utilisés ici sont des
"caractères" au sens strict. Il serait également
possible, à partir du caractère "tétrapode"
d'effectuer d'autres comparaisons plus poussées en tenant compte
des états de de caractère. Les nouveaux attributs seraient alors
"5 doigts avec pouce non opposable", "5 doigts avec
pouce opposable", "un seul doigt", "2
doigts"... on voit qu'il existe plusieurs attributs quand on
envisage des "états d'un caractère". |
 |
Comment passer du tableau de comparaison
(que l'on pourrait nommer également "tableau d'attribution")
à une classification emboîtée ?
La règle est que les regroupements ne
peuvent se faire qu'en tenant compte des attributs présents et donc
partagés ou non par les êtres vivants. Ainsi, les 6 animaux décrits
peuvent tous être regroupés dans une même "boite"
correspondant à la présence du caractère "vitellus". parce
qu'ils possèdent tous ce caractère en commun, ces animaux sont
apparentés. Si on considère le caractère "tétrapode",
celui-ci est partagé par par tous les animaux décrits sauf le thon. La
règle stipule donc que l'on peut regrouper les autres animaux dans une
boite correspondant à la présence de 4 membres mais on ne créé pas
de boite "absence de pattes". En effet, on ne peut apparenter
les êtres animaux que sur la base du partage de caractères, pas sur ce
qu'il ne partagent pas. En d'autres termes la présence de caractère
est une preuve, un fait observable indiscutable, alors que l'absence de
tel ou tel caractère ne permet aucune interprétation.
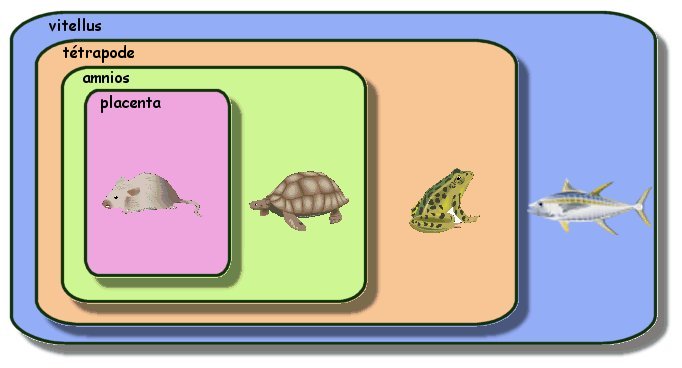 |
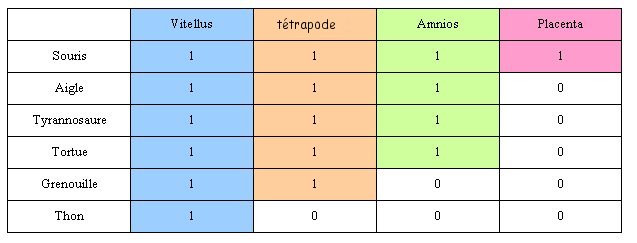
Un exemple de classification emboîtée : Tous les êtres vivants décrits dans l'image ci contre sont
inclus dans la boite (groupe) bleue : cela signifie qu'ils ont
tous en commun l'attribut "vitellus".
Si l'on considère l'attribut "tétrapode" :
souris, tortue et grenouille le possèdent. Ils sont donc inclus
dans cette boite (ou groupe) . Par contre le thon en est exclu
mais il n'a pas été créé pour autant un groupe "non
tétrapode".
|
|
Règle des parentés : plus deux êtres
vivants partagent des attributs en commun plus ils sont
apparentés. Ici la souris et la tortue partagent 3 attributs en commun
(amnios, tétrapode et vitellus), ce sont les deux animaux les
plus apparentés parmi les 4 décrits. |
Autres emboîtements possibles
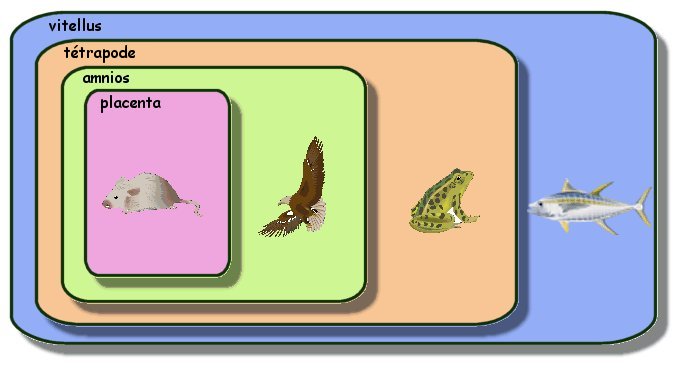 |
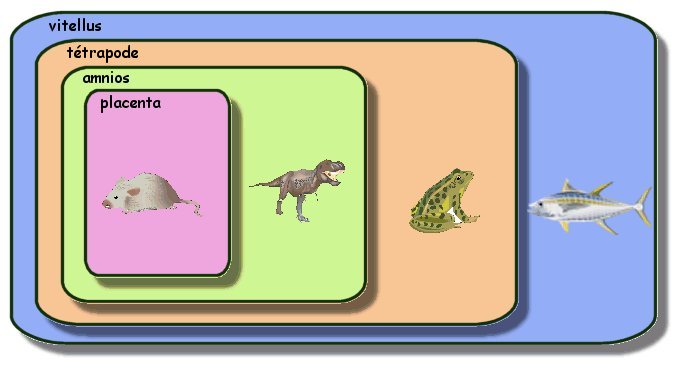 |
|
De la même façon on peut placer
l'aigle ou le tyrannosaure dans la même boite que la tortue. Il
est intéressant de constater que cette méthode permet de placer
des fossiles (tel le dinosaure) dans les boites. La méthode
consiste bien à montrer des liens de parenté et non à
reconstituer des généalogies. |
Vous
pouvez télécharger "évolcarest" pour effectuer des
classifications emboîtées : à partir du sommaire vous devez effectuer
les deux activités "classer1" et "classer2"
|
Lien
pour télécharger et installer évolcarest |

|
D'autres logiciels téléchargeables pour étudier les liens de parenté
et classer les êtres vivants :
|
Lien
pour télécharger et installer phylogène |

|
|
Lien
pour télécharger et installer phyloboite |

|
|
Lien
pour télécharger et installer phylogénia |

|
Comment passer des classifications par emboîtement à la figuration
d’arbres phylogénétiques
|
A partir de la classification emboîtée, il est
possible de fabriquer un arbre phylogénétique.
On considère que si deux êtres vivants possèdent un attribut
en commun, c'est qu'ils ont eu un ancêtre commun possédant cet
attribut. A partir de cet ancêtre commun, deux lignées ont
divergé de façon aléatoire et indépendante, pour donner deux
espèces proches.
Par exemple le tyrannosaure et la souris partagent l'attribut
"amnios" : ils ont eu un ancêtre commun (noté A3)
possédant cet attribut. Par la suite l'attribut
"placenta" (en rose) est apparu dans une seule des deux
lignées.
Graphiquement, on représentent les liens de parentés ainsi
établis par des branches. Chaque "nœud" (en rouge
foncé sur l'image) représente l'ancêtre commun aux êtres
vivants situés au dessus.
|
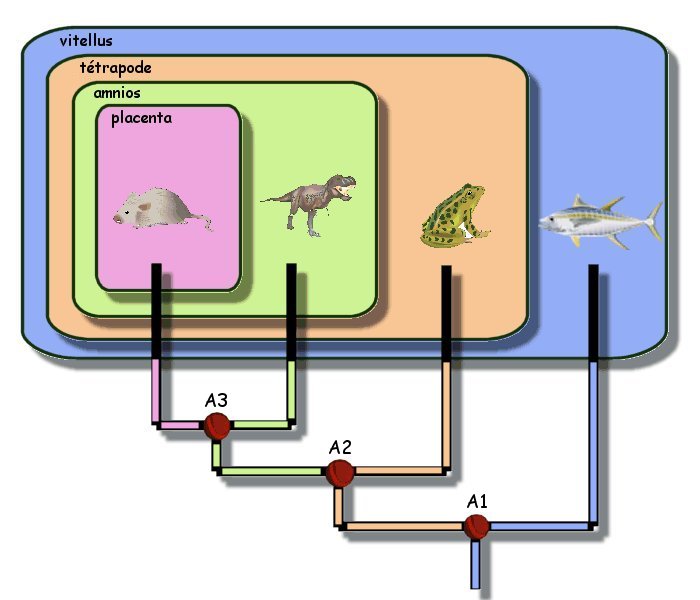 |
|
Ainsi A1 est l'ancêtre commun aux 4
animaux décrits : il possédait l'attribut "vitellus"
(en bleu) puisque cet attribut est partagé par les 4 animaux.
Dans une lignée divergente apparaît alors l'attribut
"tétrapode". cet attribut étant partagé par la
souris, le dinosaure et la grenouille, ces 3 animaux ont tous un
même ancêtre commun noté A2, possédant cet attribut. de proche
en proche on établit un arbre phylogénétique dans lequel chaque
branche symbolise un lien de parenté.
Remarque importante : Les branches de l'arbre
phylogénétique figurent des liens de parenté et non une
généalogie. L'arbre phylogénétique n'est pas un arbre
généalogique dans lequel figurent les "filiations"
d'une espèce à une autre. C'est pourquoi dans un arbre
phylogénétique tous les êtres vivants sont placés à une même
hauteur, y compris les représentants fossiles.
|
Notions de
caractères dérivés et ancestraux
Un caractère ou un état de caractère peut être qualifié de dérivé
ou bien d'ancestral.
Un caractère ancestral est un caractère
partagé par deux êtres vivants et donc qui existait chez leur ancêtre
commun. un caractère dérivé sera, par conséquent, un caractère qui
n'existe que chez l'une ou l'autre des deux espèces apparentées. La
dénomination "ancestral" ou "dérivé" pour un
caractère donné est donc relative et ne peut s'utiliser qu'en parlant
d'espèce précises. En effet, un caractère peut être considéré
comme ancestral pour certaines espèces mis dérivé pour d'autres.
En reprenant l'exemple figurant dans
l'image ci-dessus :
-
le caractère "vitellus"
peut être considéré comme un caractère ancestral pour les 4
animaux : ce caractère est partagé par ces 4 animaux et était
donc présent chez leur ancêtre commun noté A1.
-
le caractère "tétrapode"
peut alors être considéré comme un caractère dérivé de la
lignée menant aux grenouilles, dinosaures et souris, par rapport à
la lignée menant aux thons. Ce caractère n'existe pas chez leur
ancêtre commun à tous, il n'est pas ancestral, mais dérivé.
-
mais le caractère
"tétrapode" peut être considéré comme ancestral si on
exclue les poissons de l'étude : c'est en effet un caractère
commun à tous les tétrapodes et donc un caractère possédé par
l'ancêtre commun à tous les tétrapodes.
En définitive, la désignation d'un
état dérivé ou ancestral pour un caractère n'est valable que pour
une phylogénie donnée et construite. Il est également intéressant de
comprendre que seuls les états dérivés permettent l'obtention de
différents groupes et de créer des apparentements. Dans l'exemple déjà
utilisé, il aurait été impossible de créer des emboîtements
uniquement avec des caractères partagés par tous (les 4 animaux
seraient tous dans une même boite et donc impossibles à apparenter les
uns par rapport aux autres) : c'est bien la présence de caractères
dérivés qui a permis de constituer des emboîtements de plus en plus
restrictifs.
Des
phylogénies à partir de comparaisons de caractères moléculaires
Il existe des molécules communes aux êtres vivants
ayant des ancêtres communs. On cherche à regarder les différences
dans ces molécules pour établir des phylogénies moléculaires. On
parle d’ailleurs de molécules homologues s’il y a similitude des séquences
nucléotidiques et/ou polypeptidiques (au moins 20% d’homologie ou de
similitude des séquences).
Toutes les molécules ne sont pas utilisables pour établir
des liens de parenté : seules celles pouvant à la fois se " transmettre "
à la descendance et " évoluer " sont utilisables.
De ce fait, seul l’ADN est utilisable. On y adjoint la possibilité
d’utiliser les polypeptides qui, bien que non transmis directement
d’une génération à l’autre pour la plupart, sont issus de la
transcription et de la traduction des gènes de l’ADN ; il sont
des indices imparfaits des séquences nucléotidiques de l’ADN (" l’imperfection
vient de la redondance du code génétique pour certains acides aminés).
Autre limite à l’utilisation des molécules :
il faut que la molécule étudiée soit partagée par un grand nombre
d’individus ou d’espèces puisqu’il s’agit d’établir des
liens de parenté. Plus la molécule est partagée par un grand nombre,
plus les relations de parentés seront vastes.
Un exemple : établir des liens de parentés à
partir de la comparaison des séquences des globines a
des vertébrés.
L’hémoglobine est une protéine formée de quatre
sous unités appelées globines. Chaque globine correspond donc à une
chaîne précise en acides aminés (qu’il est possible de séquencer)
associée à un hème (partie non polypeptidique permettant le transport
d’une molécule de dioxygène).
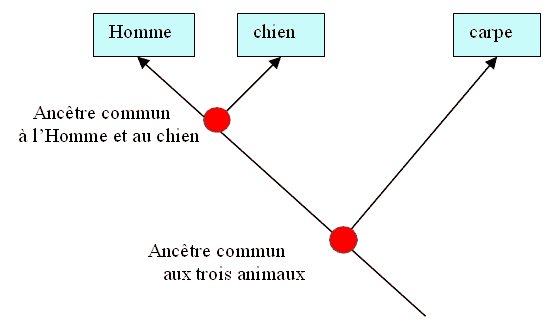
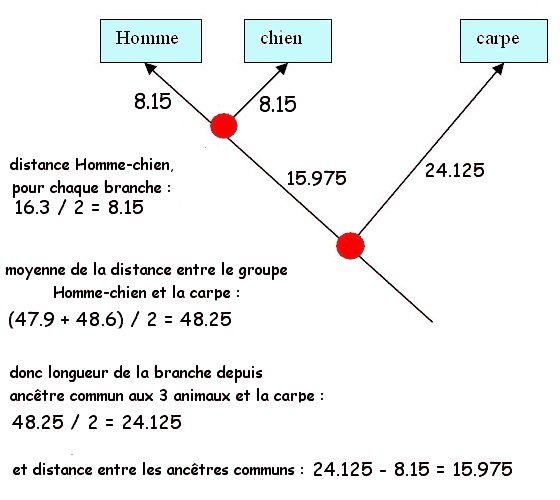
-
Comparaison Homme-chien : 16.3 % différences
-
Comparaison carpe-chien : 47.9 % différences
-
Comparaison Homme-carpe : 48.6 % différences
Les différences correspondent au nombre d’acides
aminés différents quand on compare des séquences deux à deux. Ces
valeurs peuvent être reportées dans un tableau des différences ou
tableau des dissimilitudes.
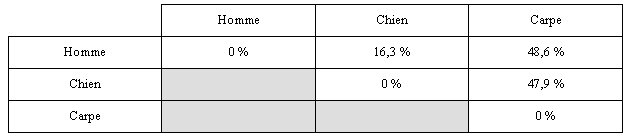
On remarque que la parenté moléculaire est plus
grande entre l'Homme et le chien : Ce sont les deux espèces dont les
globines a présentent le moins de différences.
Les Hommes et les chiens sont plus apparentés entre ex qu’avec la
carpe.
On en déduit qu’il existe un ancêtre commun à
l'Homme et au chien plus récent dans le temps que l'ancêtre commun aux
trois animaux. Les lignées Homme-chien se sont séparées plus tard que
la lignée carpe avec ancêtre commun à l'Homme et au chien.
On retranscrit alors ces informations sous la
forme d’un arbre phylogénétique :
Remarques : Si on utilise un vocabulaire rigoureux, il ne
s’agit pas d’un arbre " phylogénétique " au
sens strict mais d’un arbre " phénétique ". En
effet, cet arbre correspond à une comparaison brute des molécules sans
se poser la question des possibles convergences moléculaires dues à
des mutations reverses : en effet, même si le cas est
statistiquement rare, il est possible qu’une mutation intervienne sur
un nucléotide déjà changé et permette de retrouver la séquence
initiale.
Exemple :
ATTACGGCT devient AATACGCT par une première
mutation, puis redevient ATTACGGCT par une seconde mutation. Il y a un
retour à l’état initial ou ancestral de la séquence.
De plus, à cause de la redondance du code génétique,
certaines mutations de l’ADN ne se répercutent pas dans la séquence
du polypeptide traduit (on parle de mutations silencieuses) : dans
ce cas, la comparaison de séquences polypeptidiques peut (et c’est fréquent)
cacher certaines évolutions. Il est donc préférable de comparer des séquences
nucléotidiques plutôt que des séquences polypeptidiques.
La comparaison des molécules permet d’accéder à
des valeurs chiffrées (ce qui n’est pas le cas des comparaisons au
niveau des organes qui sont davantage " qualitatives "
avec des résultats en terme de présence et/ou d’absence de tel ou
tel caractère homologue).
Il est donc possible de tenir compte de ces valeurs
pour tracer des arbres dont la longueur des branches (qui représentent
les liens de parenté) indiquera une distance génétique : plus la
branche est longue, plus la distance est grande.
Des recherches avec des molécules peu évoluées
(ADN ribosomal, cytochrome C : pigment respiratoire) on permis de
remonter la phylogénie jusqu'aux bactéries. C'est l'association des différentes méthodes qui
permet d'établir des phylogénies les plus proches possibles de la réalité
possible de l'évolution.
|