La transmission DES CARACTERES
La plupart des élèves ont vision Lamarckienne de
l'évolution. Quand on leur demande d'expliquer comment un long cou est
apparu chez les girafes, ils répondent que celles-ci se sont adaptées
en allant chercher les feuilles de plus en plus haut dans les arbres
(concept ancien de "la fonction créé l'organe"). Ils
considèrent donc que des caractères acquis durant la vie, par
adaptabilité à un type d'environnement, se transmettent à la
descendance. Ils sont donc dans l'idée qu'il existe une
"hérédité des caractères acquis".
Deux arguments peuvent perturber cette conception de
l'évolution :
-
tout d'abord une question : un talent se transmet-il
? un virtuose du violon aura-t-il des enfants obligatoirement
virtuoses à leurs tour ?
-
ensuite la compréhension des modalités de la
transmission de l'information génétique
 Une
transmission de cellules sexuelles haploïdes Une
transmission de cellules sexuelles haploïdes
 |
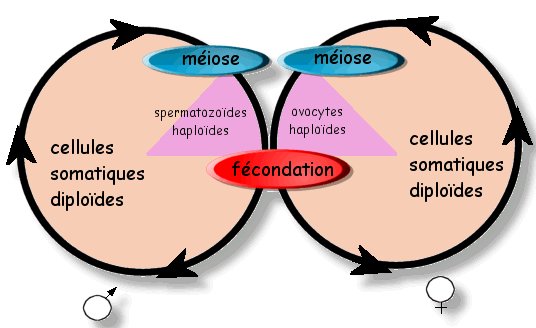
On sait que la reproduction sexuée comporte une
phase d'union de deux cellules appelée fécondation. Or une
espèce se distingue par son matériel génétique et, en l'occurrence,
son caryotype : chaque espèce a un caryotype précis qui ne peut
différer entre deux individus que s'ils sont de sexe opposé
(différence entre les chromosomes X et Y par exemple).
Une espèce diploïdes est ainsi nommée si les individus de
cette espèce sont constitués majoritairement de cellules
diploïdes, c'est à dire comportant des paires de chromosomes
homologues. C'est le cas de l'espèce Humaine et de tous les
animaux. Il existe des espèces haploïdes qui présentent, à
l'inverse, un cycle de vie dont les individus sont constitués de
cellules ayant des chromosomes en uniques exemplaires.
|
|
Prenons le cas de l'espèce humaine
caractérisée par des cellules somatiques ("soma" =
"corps") diploïdes et un caryotype avec 23 paires de
chromosomes (caryotype de gauche ci-dessus). Un problème se pose
: si deux cellules diploïdes (à 23 paires de chromosomes)
mettent en commun leur matériel génétique lors de la
fécondation, alors la cellule oeuf issue de cette fécondation
comportera deus fois 23 paires de chromosomes. la cellule oeuf n'a
plus le caryotype de l'espèce. Il faut donc envisager un
mécanisme cellulaire permettant de réduire le nombre de
chromosomes des cellules servant à la reproduction sexuée des
espèces diploïdes. Ce mécanisme existe puisqu'il est maintenant
facile de constater que les cellules sexuelles (gamètes) des
espèces diploïdes ont un caryotype comportant des chromosomes en
simple exemplaire (caryotype de droite sur l'image ci-dessus).
|
 |
Un organisme diploïde est donc constitué de
cellules somatiques diploïdes. Toutefois un tel individus (quand
il arrive à maturité sexuelle) fabrique, en nombre
limité, des gamètes haploïdes à partir de cellules
diploïdes. Le mécanisme permettant de transformer des
cellules comportant des paires de chromosomes en cellules
contenant des chromosomes en simple exemplaire se nomme la
méiose.
La méiose est un ensemble de deux divisions
cellulaires successives précédé d'une seule réplication de
l'ADN. Elle aboutit à la formation de quatre cellules haploïdes
à partir d'une seule cellule diploïde.
chez l'Homme, la méiose ne touche que les
cellules dites germinales (qui sont diploïdes) afin de les
transformer en cellules sexuelles (spermatozoïdes ou ovocytes
selon le sexe). Ainsi la fécondation s'effectue entre des
cellules haploïdes et aboutit donc à la formation d'une cellule
oeuf diploïde, rétablissant par là même le caryotype de
l'espèce.
Le cycle de développement de toutes les
espèces à reproduction sexuée est marqué par l'alternance
d'une phase diploïde et d'une phase haploïde.
|
-
La méiose est donc un mécanisme cellulaire
permettant le passage de la diploïdie à l'haploïdie
-
La fécondation de deux gamètes haploïdes rétablit
la diploïdie
-
Les deux phénomènes sont interdépendant dans un
cycle de vie d'une espèce à reproduction sexuée, l'association
des deux phénomènes permettant ainsi une stabilité des caryotypes
des espèces.
La méiose :
une double division cellulaire permettant le passage à l'haploïdie
 |
Le schéma ci-contre montre sommairement quelques
phases de la méiose. On remarque que la méiose est une succession de deux divisions
cellulaires précédées d'une seule phase de réplication (ou
duplication) de l'ADN.
Ainsi, à partir d'une cellule diploïde, comportant des paires
de chromosomes à deux chromatides :
-
une première division va séparer la cellule initiale en
deux cellules comportant des chromosomes en simple exemplaires
mais toujours à deux chromatides (séparation des chromosomes
homologues en deux lots égaux)
-
une seconde division va séparer les chromatides de chaque
chromosome en deux lots égaux.
-
il y a donc, au final, fabrication de 4 cellules haploïdes
contenant des chromosomes en simple exemplaire et à une
chromatide
|
Réponse au
problème de l'hérédité des caractères acquis
On comprend que l'innovation génétique (qui a pour
origine des mutations de l'ADN) ne peut se transmettre à la descendance
que via des cellules sexuelles. Ces cellules sexuelles n'ont évidemment
aucun lien direct avec l'environnement (en revenant à nos
girafes, il est difficile de s'imaginer qu'une utilisation fréquente du
cou puisse modifier le contenu de l'information génétique des
gamètes).
De même, des mutations affectant les cellules germinales
(à l'origine des gamètes) ont la possibilité d'être transmises à la
descendance mais n'auront aucune chance d'avoir des conséquences sur le
phénotype du porteur de ces gamètes (sauf s'il s'agit d'une mutation
cancérigène).
Donc, seules les mutations affectant les cellules
germinales (et non somatiques) ont des chances de devenir héréditaires
et ainsi de "rentrer" dans l'évolution des populations. En
effet, un individu issu d'un gamète comportant une mutation
particulière aura toutes ses cellules (y compris somatiques) porteuses
de cette innovation (toutes ses cellules sont issues des divisions à l'identique
de la cellule oeuf originelle). cela implique que cet individu, quand il
fabriquera à son tour ses propres cellules sexuelles, elle seront
toutes porteuse de la mutation. Ainsi l'innovation génétique se répand
dans la population, génération après génération.
La
disjonction des allèles
Si la méiose peut être considérée comme un mécanisme
participant à la stabilité de l'espèce parce qu'elle permet le
maintien des caryotype (en association avec la fécondation), on peut
également considérer qu'elle à l'origine d'une certaines variabilité
génétique.
En effet lors des différentes phase de la méiose, des
chromosomes homologues vont être séparés. Ces chromosomes homologues
(d'une même paire) comportent bien les mêmes gènes mais ils sont
issus de deux gamètes différents à l'origine : un des deux
chromosomes d'une même paire à pour origine un gamète paternel et
l'autre est d'origine maternelle. Ainsi les gènes des chromosomes
homologues sont les mêmes mais peuvent présenter des versions
différentes appelées allèles.
La séparation des chromosomes homologues (en première division
de méiose) s'effectue de manière aléatoire entre les chromosomes de
paires différentes.
Par exemple :
-
le chromosome 1 paternel (1p) sera obligatoirement
séparé du chromosome 1 maternel (1m)
-
le chromosome 2 paternel (2p) sera obligatoirement
séparé du chromosome 2 maternel (2m)
-
mais 1p peut aussi bien se retrouver dans un lot
avec 2p aussi bien qu'avec 2m : cette répartition aléatoire
des chromosomes non homologues est appelé brassage
interchromosomique.
De plus, en début de méiose, les chromosomes homologues
s'apparient (réunion par paires d'homologues). Lors de cet appariements,
les chromatides de ces chromosomes se croisent et peuvent s'intervertir
(on parle de "crossing over"), créant ainsi des nouveaux
chromosomes à la fois porteurs d'informations d'origine paternel et
d'origine maternelle. On parle alors de brassage intrachromosomique.
Ces brassages des chromosomes ne fabriquent pas
d'innovation génétique au sens strict : il n'y a en effet pas de
nouveaux gènes ou allèles créés par un tel brassage. Par contre ces
brassages permettent l'apparition de nouvelles associations d'allèles,
dans les gamètes, qui n'existaient pas chez les parents. Ainsi chaque
gamète est différent du gamète voisin (cela explique par ailleurs
pourquoi des faux-jumeaux, issus de gamètes différents, ne se
ressemblent pas : les nouvelles associations d'allèles sont multiples :
une chance sur 300 millions que les deux spermatozoïdes à l'origine
des deux enfants présentent la même association d'allèles).
Pour conclure, le brassage génétique lors de la méiose
est donc à l'origine d'une variabilité des génomes. On peut
également dire que la fécondation va amplifier cette variabilité en
réunissant au hasard deux gamètes issus d'individus différents.
|
Télécharger une application présentant le double
brassage des chromosomes lors de la méiose |
|
|