|
||||||
Des génotypesL'acide désoxyribonucléique, l'ADN, est une macromolécule porteuse du programme génétique. Une partie seulement de cette molécule sert directement à la fabrication des protéines, réalisant ainsi les phénotypes à différentes échelles. Ces parties de l'ADN sont dites codantes et sont également appelées "gènes" (Par ailleurs le mot "gène" fait référence au verbe "générer"). Le génotype correspond à l'ensemble des gènes, portés par les chromosomes, d'un organisme. Un gène se définit autant par la protéine qu'il fabrique que par sa position sur un chromosome (locus du gène). D'un être vivant à l'autre le génotype peut présenter des différences :
Il est important de bien comprendre la relation entre le génotype et le phénotype : si c'est bien ce dernier qui est soumis à la sélection naturelle (et donc au filtre de l'évolution), ce sont bien les gènes qui seront soumis à des variations (mutations) et donc qui seront à la base de l'innovation.
L'ADN se trouve dans le noyau des cellules eucaryotes et des protéines se trouvent dans le cytoplasme : l'information génétique est donc localisée dans le noyau alors que les protéines doivent pouvoir se retrouver à tout endroit de la cellule. L'ADN est une macromolécule de plusieurs millions de nucléotides alors qu'une protéine est constituée de quelques dizaines, voire centaines, d'acides aminés : une même molécule d'ADN comporte plusieurs gènes, chacun pouvant être à l'origine d'une protéine. Il doit exister un intermédiaire entre un gène de l'ADN et la protéine dont il est à l'origine.
Au cours du cycle cellulaire, dans le noyau, un gène de l'ADN est transcrit en une molécule complémentaire : l'ARN messager (ARNm) Une partie de la molécule d'ADN s'ouvre (car double chaîne enroulée en hélice) sous l'action d'enzymes spécifiques (ARN polymérase) et un seul des deux brins va servir de modèle : on parle de brin transcrit de l'ADN. La molécule d'ARNm va être fabriquée à partir du brin transcrit. Tout l'ADN n'est donc pas transcrit, mais uniquement une partie correspondant à un gène. La transcription de l'ADN en ARNm suit le principe de la complémentarité des bases avec une exception : l'uracile remplace la thymine sur l'ARNm. La molécule d'ARNm reflète donc une partie du programme génétique. Elle a la propriété de sortir du noyau par les pores nucléaires pour se rendre dans le cytoplasme.
L'ARNm comporte une séquence de nucléotides avec les bases azotées suivantes : adénine, guanine, cytosine et uracile. Une protéine correspond à un enchaînement d'acides aminés reliés entre eux par des liaisons peptidiques. La succession de ces acides aminés n'est pas le fruit du hasard : de cette succession (nombre + ordre) dépend la nature et les propriétés de la protéine. La traduction de la séquence nucléotidique de l'ARNm en séquence d'acides aminés du polypeptide suit un code génétique : à chaque triplet (ou codon) de nucléotides de l'ARNm correspond un acide aminé.Remarque :
La traduction de l'ARNm en protéine se fait dans le cytoplasme. Elle est assurée par les ribosomes (u n ribosome est constitué par de l'ARN ribosomial et des protéines, répartis en deux sous-unités, l'une servant de cadre de lecture et de fixation à l'ARNm, l'autre servant à la mise en place des acides aminés). D'autres molécules interviennent aussi :
La traduction de l'ARNm commence au niveau d'un codon spécifique dit initiateur : AUG. Il permet la mise en place d'une méthionine. L'élongation de la chaîne polypeptidique se fait en respectant la séquence de l'ARNm, codon par codon. La traduction se termine quand le ribosome arrive sur un codon stop (UGA, UAA, UAG) pour lequel ne correspond aucun acide aminé.
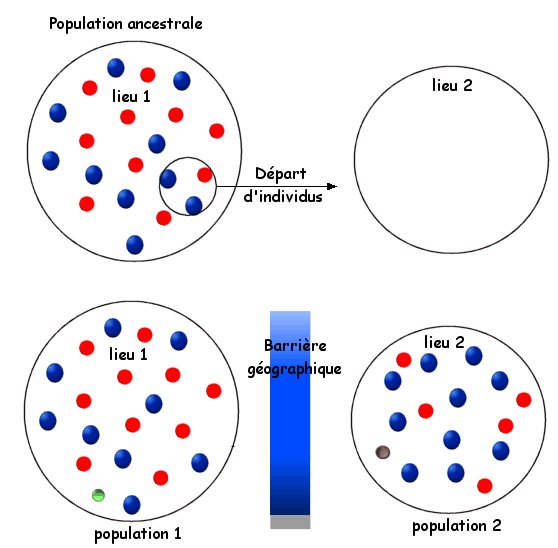
Au niveau individuel, un génotype peut-être soumis indirectement (via le phénotype exprimé) à la sélection naturelle. Les conséquences seront bien évidemment également individuelles : le génotype sera soit avantageux, soit neutre, soit défavorable à l'individu. Darwin aurait parlé de génotype adapté (ou non) compétitif (ou non) s'il avait eu connaissance des gènes. Un génotype "avantageux" se reproduira plus et se retrouvera à forte fréquence au bout de plusieurs générations si la pression de sélection ne varie pas. On voit bien que ce n'est pas l'individu qui évolue. Les mutations permettent la variation des séquences géniques, faisant apparaître, au hasard, de nouvelles versions de ces gènes (on parle de polyallélisme quand un gène supporte plusieurs variantes ou allèles). Il y a donc au sein d'une même espèce plusieurs individus dont les génotypes sont différents par les différents allèles que chacun possède. On constate par l'observation et l'analyse des génotypes que certains allèles sont plus fréquents dans certaines populations géographiques que dans d'autres, alors que l'ensemble des individus de ces populations sont de la même espèce. Une population ne se définit donc pas par des gènes particuliers mais par des fréquences des allèles de ces gènes. Comment expliquer la différence de fréquence des allèles entre deux populations d'une même espèce alors que les mutations sont aléatoires et que les deux populations sont sensées être issues d'une même population ancestrale ?
Cet exemple illustre ce que l'on appelle la dérive génétique des populations ou microévolution. La microévolution est donc une variation des fréquences alléliques se produisant dans le patrimoine génétique, et donc l'ensemble des génotypes de l'espèce, de génération en génération. Pour ces raisons, les généticiens considèrent que les individus représentent une distribution de génotypes mais que c'est bien la population qui engendre (au sens "transmission") ces génotypes. Il en résulte que l'unité génétique en évolution, c'est la population, avec un ensemble de génotypes et non l'individu avec un génotype unique. Remarque : La dérive génétique, ou microévolution, est le concept central de la théorie synthétique de l'évolution. S'il est difficile de ne pas souscrire à ce mécanisme de l'évolution il faut considérer qu'il n'est pas le seul. En effet, la microévolution n'explique que les cas d'évolution graduelle avec des populations évoluant très lentement dans le temps jusqu'à la spéciation. La spéciation est associée au terme de macroévolution. Les observations de fossiles montrent que l'évolution n'est pas toujours graduelle mais que des cas de macroévolution se sont opérés sur de courts intervalles de temps : c'est pour cette raison que la théorie synthétique de l'évolution s'est vue enrichie par la théorie des équilibres ponctués (S. J. Gould) qui envisage des vitesses dévolution différentes (Gould propose des durées longues pendant lesquelles les espèces n'évoluent pas ou peu, juste une microévolution envisagée, ponctuées par des périodes courtes présentant des macroévolutions).
|
||||||