Sélection naturelle
 Un petit historique Un petit historique
 La sélection naturelle selon Darwin (1809-1882)
La sélection naturelle selon Darwin (1809-1882)
La " sélection naturelle " permet
à Darwin d’expliquer comment les espèces se différencient au cours des
temps géologiques. Les fondements de cette théorie reposent sur ses
observations au cours d’un tour du monde de 5 ans à bord du
Beagle : l’étude de différentes espèces de pinsons des îles Galapagos montrait des adaptations de ces oiseaux à des environnements
particuliers. Il publiera le célèbre " De l’origine des
espèces… " en 1859 soit 23 ans après son voyage.
En premier lieu Darwin remarque que la reproduction
au sein des espèces devrait amener à une croissance exponentielle des
individus. Pourtant les populations restent stables. A cela s’ajoute
que les ressources naturelles sont limitées et plus ou moins constantes
sur le long terme (abstraction des phénomènes saisonniers cycliques).
Se basant sur un concept de Malthus (économiste qui a rédigé des
essais sur la croissance des populations humaines) Darwin en déduit qu’il
y donc un équilibre qui s’installe entre les populations et les
ressources naturelles disponibles. Une nouvelle déduction découle de
la première : une concurrence s’installe donc entre les êtres
vivants.
Un autre fait se rajoute à cette dernière
déduction : dans une même population les individus sont différents
(on parle aujourd’hui de variabilité allélique ou polymorphisme des
espèces). Il en déduit qu’une population comporte donc des individus
potentiellement plus adaptés que d’autres et que la mise en
concurrence de ceux-ci amènera à sélectionner les plus aptes. Il y a
donc une "sélection naturelle" qui va aboutir à une
transformation graduelle des populations en sélectionnant des
caractères favorables. La suite de ces transformations amenant à une
spéciation, par "persistance des plus aptes".
Darwin pondère son propos en ajoutant que tous les
caractères ne sont pas pris en compte par une sélection naturelle :
certains caractères sont favorables, d’autres nuisibles mais la
plupart restent "insignifiants" (pas de prise de la sélection
naturelle sur ces caractères qui vont donc persister). Les caractères
sélectionnés vont alors se transmettre de façon héréditaire.
Les principales nouveautés portent donc surtout sur
une théorisation des causes évolutives , "la sélection
naturelle", pour laquelle il apporte nombre de faits d’observations
issues de ses voyages. Il se démarque aussi de Lamarck en considérant
la possibilité de culs-de-sac évolutifs dans lesquels on voit
disparaître des populations ou espèces que la sélection naturelle
élimine.
La théorie neutraliste
Motoo Kimura (1924-1994) complétera avec la
" théorie neutraliste " avançant que les mutations
aléatoires ne sont pas forcément soumises à la sélection naturelle
et que l’évolution peut aussi être le fruit du hasard : en cela
il propose une évolution non guidée, s’écartant de tout finalisme.
Il y a donc une dérive génétique aléatoire expliquant le fort
polymorphisme des populations.
Qu’est-ce qui est le plus couramment admis aujourd’hui ?
On ne peut plus considérer la " sélection
naturelle " comme une cause véritable de l’évolution, mais
certains mécanismes décrits par Darwin restent tout à fait
scientifiquement recevables (à la lumière des connaissances
actuelles).
Darwin se trompe quand il dit que l’environnement
" oblige " les organismes à s’adapter et donc
créé de l’innovation : il reste dans l’idée que
" la fonction créé l’organe " de Lamarck et
pense également que les adaptations réussies par des organismes au
cours de leur existence se transmettent à la descendance (ce qui
paraît aujourd’hui non recevable puisque les découvertes en
génétique ont montré que, globalement, il ne doit pas y avoir de
transmission héréditaire de caractères acquis).
Darwin a raison de donner un rôle à l’environnement
dans la sélection de certains caractères : il y a manifestement
des formes de vie plus adaptées que d’autres dans un environnement
donné. Par contre la question était de savoir si l’environnement
avait le " pouvoir " de transformer les êtres
vivants.
Enfin, mais Darwin le disait déjà, toutes les
attributions ne sont pas soumises à sélection : des formes ne
présentent ni avantage, ni inconvénient dans un environnement donné,
on dirait aujourd’hui que la majorité des allèles (issus des
mutations) sont neutres, ce qui explique la grande diversité intraspécifique. Si toute caractéristique était soumise à une
sélection naturelle, les formes de vie auraient une tendance à toutes
se ressembler et on n’observerait pas l’actuelle diversité.
La sélection agit donc, à la manière d’un
filtre, sur quelques caractères dont l’existence initiale repose
totalement sur le hasard des mutations qui les ont fait apparaître. La
" sélection naturelle " n’est donc pas la cause
de l’évolution mais le tamis sélectif réduisant la diversité
possible des formes de vie issues du hasard et des aléas de la
génétique.
Un exemple
théorique
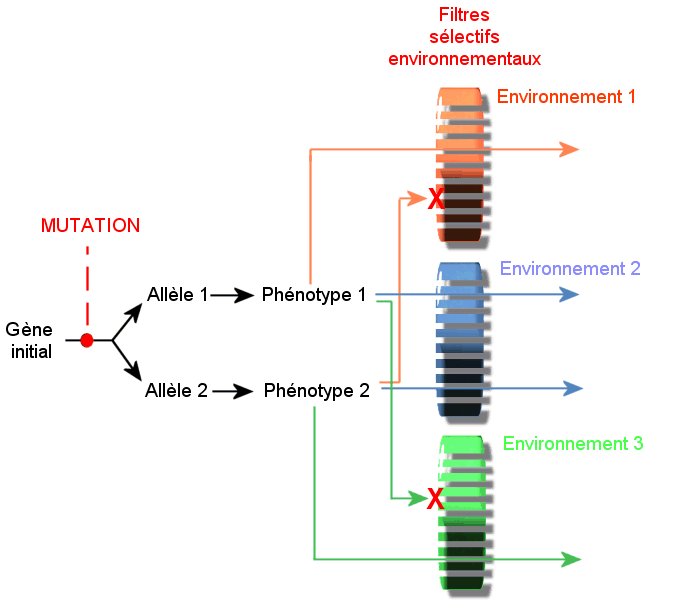
Une mutation affecte un gène, faisant apparaître
ainsi deux allèles (versions différentes) de ce gène. Chaque allèle
est à l’origine de deux phénotypes différents. Il y a donc eu une innovation génétique, causée au
hasard par une mutation qui a pour conséquence une augmentation de la
diversité.
Cette diversité est observable mais elle diffère
selon les environnements. Il y a donc un filtre.
Si les deux phénotypes sont placés dans un
environnement 1, défavorable au phénotype 2 : au bout de
plusieurs générations, seul le phénotype 1 s’observera dans cet
environnement. La mutation n’ est donc pas neutre.
Si les deux phénotypes sont placés dans un
environnement 2, ni défavorable, ni favorable pour les deux
phénotypes : au bout de plusieurs générations, les deux
phénotypes perdurent. La mutation est neutre.
Si les deux phénotypes sont placés dans un
environnement 3, défavorable au phénotype 1 : au bout de
plusieurs générations, seul le phénotype 2 s’observera dans cet
environnement. La mutation n’ est donc pas neutre.
Comme on le voit une mutation n’est pas favorable
ou défavorable, à priori. Cela montre bien l’aspect aléatoire de la
mutation, non seulement en tant que cause mais également dans ses
conséquences : ce sont les mutations qui sont à l’origine de la
diversité. Cela montre que l’environnement ne provoque pas une
évolution des formes de vie mais exerce une pression sélective qui
peut aboutir à une diminution de la diversité (environnements 1 et 3
de l’exemple) ou bien à son maintien (environnement 2).
Cet aspect du rôle de l’environnement est d’ailleurs
subtilement exploité par les néo-créationnistes : le filtre
environnemental, en sélectionnant certains phénotypes plutôt que d’autres,
donne l’illusion d’une évolution guidée, dirigée et qu’il
existe donc une forme de finalisme (si finalité il y a, alors elle est
d’essence divine).
Il n’y a aucune finalité, on voit bien qu’il y a
tout d’abord un hasard (c’est ce hasard que les créationnistes
refusent le plus, en fait), puis une sélection non guidée : l’évolution
est hasardeuse et opportuniste, elle n’obéit à aucune
" feuille de route ".
Trois exemples de pression sélective environnementale
La
Phalène du Bouleau
|
La phalène du bouleau (Biston
betularia L.) est un papillon nocturne des régions tempérées. Ce papillon se rencontre sous
deux formes, une de couleur claire dit typica et l'autre
sombre dit carbonaria ou mélanique (la couleur des
ailes étant due à la quantité d'un pigment appelé
mélanine présent dans les cellules). Le déterminisme génétique
de cette coloration est monogénique et autosomique, l'allèle
carbonaria c+ étant dominant sur l'allèle typica c [forme mélanique] :
cc+ ou c+c+ [forme claire] : cc.
|
 |
|
Forme claire et sombre de la Phalène du Bouleau |
|
Au cours du XIXe siècle,
des biologistes anglais observent que la forme sombre devient plus fréquente
à proximité des villes industrielles (observée pour la première fois
en 1848 dans la région de Manchester, cette forme sombre est devenue
largement majoritaire en 1895 dans cette même région : 98%
de la population). Ce phénomène est à mettre en relation avec
la pollution atmosphérique issue des résidus de combustion du
charbon (émis par l'industrie locale). En conséquence, les
troncs et les branches des arbres devenaient plus sombres (à la fois
par les dépôts de fumée mais surtout par la disparition des lichens plus
clairs qui les recouvraient).
Ces papillons nocturnes se posant en journée sur les arbres, il a
été montré que le taux de survie des individus de type sombre était
plus élevé que celui des individus de type clair, probablement parce
que ces derniers étaient plus visibles aux yeux de leurs prédateurs
oiseaux.
|

|
|
Formes claire et sombre sur tronc
"pollué" |
|
Vers la fin des années 1960, des mesures sont prises
pour améliorer la qualité de l'air. La diminution de la pollution
provoque un retour à des troncs d'arbres plus clairs (moins de
suie déposée sur les écorces, retour des lichens). On observe
parallèlement un retour des formes claires de phalènes.
|

|
|
Formes sombre et claire sur tronc
"propre" |
Les évolutions rapides constatées au sein des populations des phalènes
du bouleau, mises en relation avec des facteurs environnementaux
et une pression de prédation, représentent un exemple souvent utilisé
pour illustrer la sélection naturelle. Cet exemple peut être
partiellement remis en cause pour deux raisons principales :
-
le mélanisme intervient également dans la thermorégulation de
ces insectes : ce n'est donc pas qu'un facteur de camouflage
-
les premières études sont statistiquement discutables (méthodes
de comptage à revoir)
Mais :
-
il a été constaté le même phénomène d'évolution des
populations chez d'autres insectes
-
une étude récente est en cours, tenant compte de meilleures
méthodes de comptage : elles semblent appuyer l'idée d'une
sélection naturelle rapide par pression de prédation.
Drépanocytose et environnement à paludisme
La drépanocytose est une maladie due à la présence
d'hémoglobine anormale dans les globules rouges. C'est une mutation
ponctuelle du gène codant pour la chaîne b
de l'hémoglobine qui provoque l'apparition de l'allèle HbS (HbA étant
l’allèle normal).
Les humains étant des organismes diploïdes, ils ont
tous leurs chromosomes en double exemplaire. En conséquence, chacune de
nos cellules possède deux fois le gène permettant la fabrication de la
chaîne b de l'hémoglobine. Par contre la
version de ce gène, l’allèle, n’est pas obligatoirement la même d’un
chromosome à l’autre, donc trois cas sont possibles :
-
Une personne avec les deux allèles
" sains " : cette personne est normale et
transmet toujours un allèle HbA à sa descendance.
-
Une personne avec l’allèle HbA et l’allèle HbS : cette
personne est " porteuse saine ", c'est-à-dire
qu’elle ne développe pas la maladie mais est susceptible de
transmettre l’allèle HbS à sa descendance (un risque sur deux)
-
Une personne avec les deux allèles HbS sera malade et la maladie
est le plus souvent mortelle avant la puberté.
Statistiquement, la drépanocytose étant une maladie
mortelle avant que les porteurs malades ne puissent se reproduire, l’allèle
HbS devrait disparaître avec le temps, la mutation étant clairement
défavorable. En fait, elle ne devrait même plus exister. Pourtant
cette maladie, qui cause des ravages sur le continent africain
principalement, perdure.

On peut donc envisager que la mutation est
défavorisante dans certains environnements mais qu’elle peut aussi
conférer un avantage dans d’autres environnements.
On remarque que la drépanocytose est maintenue dans
les régions où sévit le paludisme, maladie qui n’a rien d’héréditaire
puisqu’il s’agit d’une maladie parasitaire transmise par l’anophèle.
Quel lien entre drépanocytose et paludisme ?
Les individus homozygotes porteurs de l’allèle HbS
ne sont pas plus avantagés dans un environnement à paludisme que dans
un environnement dépourvu de ce fléau : la maladie est mortelle.
Par contre il a été constaté que les individus hétérozygotes pour
le gène de la béta-globine (individus porteurs à la fois de l’allèle
HbA et HbS) sont davantage protégés contre le paludisme que les
individus homozygotes " sains ", porteurs de l’allèle
HbA uniquement. Il en résulte dans les zones à paludisme :
-
Une mortalité des homozygotes HbS // HbS (pression de sélection
négative de la mutation)
-
Une plus forte mortalité des homozygotes HbA // HbA que des
hétérozygotes HbA // HbS (pression de sélection négative du
paludisme compensée par une pression de sélection positive de la
mutation pour les hétérozygotes)
-
Donc l’allèle HbS est maintenu dans les populations vivant en
zone de paludisme.
Comme on le voit la même mutation a des
conséquences différentes ou convergentes selon les
environnements : on comprend ainsi un des mécanismes expliquant qu’à
partir d’une population originelle, on aboutit à deux populations
(séparées géographiquement) présentant des fréquences d’allèles
différentes.
Le sixième doigt du Panda
Le doigt supplémentaire, ou
"faux pouce", formé par un os du carpe (l'os
sésamoïde radial) hypertrophié est d'une importance vitale pour
le célèbre panda géant et son petit semblable (sans parenté
directe), le panda rouge, qui l'utilisent pour attraper plus
facilement les pousses de bambous dont ils se nourrissent.
|

|
 |
|
Panda roux |
Panda géant |
En l'observant, le paléontologue américain Stephen Jay Gould avait
élaboré, il y a plus d'un quart de siècle, son concept de contingence
dans l'évolution, c'est-à-dire une hypothèse selon laquelle la vie
est soumise non pas à des transformations lentes et continuelles, comme
l'imaginait Darwin, mais à des choix rapides et aléatoires.
Le faux doigt du panda n'est pas apparu progressivement mais de
façon brutale et n'est donc pas la conséquence d'une adaptation à un
régime alimentaire.
Par contre il est indéniable que ce sixième doigt facilite
l'alimentation des pandas et donc qu'une pression de sélection positive
s'exerce chez les mangeurs de bambou pourvus de cet appendice
supplémentaire. Nous ne sommes pas dans le célèbre (mais faux)
"la fonction créé l'organe" de Lamarck mais plutôt dans un
"l'organe peut améliorer la fonction". Contrairement à ce
que les paléontologues supposaient, le fameux sixième doigt ne
lui servait pas à l'origine à manipuler de la nourriture, mais
à faciliter ses déplacements. En effet, la découverte
d'un ancêtre du panda roux, appelé Simocyon batalleri, montre
que cet animal vieux de neuf millions d'années possédait, tout
comme son descendant herbivore, un sixième doigt.
En étudiant sa denture, il a été découvert
que cet animal mangeait essentiellement de la viande, et non des
végétaux comme le petit panda actuel. C'est pourquoi Simocyon n'utilisait pas son sixième doigt pour saisir les
pousses de bambou comme le fait aujourd'hui le petit panda, mais
plus certainement pour aider à sa locomotion dans les arbres
: le squelette de Simocyon, de la taille d'un puma,
présente des particularités morphologiques qui indiquent un mode
de vie arboricole. Les données recueillies indiquent qu'il vivait
dans un environnement peuplé de nombreux prédateurs. Dans ce
contexte ce carnivore plutôt charognard et peu véloce
pouvait donc leur échapper en grimpant aisément dans les arbres.
|

Simocyon batalleri |
Aujourd'hui, des millions d'années ont passé ; l'environnement et le
mode de vie du petit panda sont très différents de ceux de Simocyon, au
point de rendre obsolète l'utilisation originelle de son sixième doigt.
Mais comme son régime alimentaire s'est aussi modifié, l'usage premier
de ce faux pouce s'est transformé en un usage second et unique, à savoir
la manipulation de bambous et autres feuillages. Le faux pouce du
petit panda est un merveilleux exemple d'exaptation, ou réadaptation
secondaire, soit une structure dont la fonction ou l'usage actuel est
différent de la fonction originelle.
En
conclusion
-
le sixième pouce du Panda est apparu de façon brutale et
aléatoire (on parle de contingence) sans être la conséquence
d'une sélection naturelle.
-
sa fonction première a été de faciliter les déplacements de
l'animal primitif dans les arbres : on peut dire ici que l'environnement
a sélectionné favorablement cette innovation.
-
au cours des temps géologiques, l'alimentation des Pandas a
changé, ils sont devenus végétariens en mangeant de façon
exclusive des pousses de bambou. La fonction du sixième doigt a
également changé, devenant un "outil" intéressant pour
ce régime alimentaire.
La sélection naturelle n'est pas source d'innovation mais peut
favoriser (ou l'inverse) et maintenir un caractère particulier sans
pour autant lui attribuer un seul intérêt : l'évolution est
opportuniste.
|